¶ Authors
Michelle W. Wu1,2,3, Nazim Kourdougli1, and Carlos Portera-Cailliau1,4
1 Department of Neurology, David Geffen School of Medicine at the University of California Los Angeles
2 Neuroscience interdepartmental graduate program, University of California Los Angeles
3 UCLA-Caltech Medical Scientist Training Program
4 Department of Neurobiology, David Geffen School of Medicine at the University of California Los Angeles
¶ Content
- Nomenclature and other challenges in the field
- Neuronal domains in vitro reflect correlated calcium signals, but not synaptic activity
- Early network oscillations revealed by calcium imaging in acute brain slices
- ENOs in vivo
- Interneurons also participate in synchronous network events
- Spindle bursts revealed by electrophysiology
- Activity patterns in embryonic cortex
- Developmental sequence of events
¶ 1. Nomenclature and other challenges in the field
One of the challenges everyone faces when approaching this topic is making sense of the terminology that is used to describe correlated network activity (see Network event terminology). The large number of terms (and alphabet soup of corresponding abbreviations) used to describe different patterns of synchronous activity can give rise to unnecessary confusion. This is in part because studies that investigated the subject used different experimental preparations (acute brain slices vs. in vivo recordings in the intact brain) and different recording techniques (electrophysiology vs. calcium imaging), and also focused on different brain regions or animal species. As a result, different groups of investigators chose different terms to describe activity phenomena that were potentially similar or even identical. Here, we make a concerted effort to simplify and clarify the matter. First, we focus on a single brain region, the neocortex, and on events that take place in early postnatal rodents. Although much pioneering research on correlated network activity was carried out in the hippocampus, there are important differences between activity in both regions. Second, we incorporate results of different studies into a single, cohesive narrative about developmental changes in network activity. This requires some interpretation of the available data and, inevitably, some readers may disagree with our take, which will hopefully lead to a fun debate. Third, we propose a new term to refer to synchronous neuronal activity during development: correlated network activity (CNA).
Table: Historical Timeline of studies on neonatal spontaneous activity
¶ 2. Neuronal domains in vitro reflect correlated calcium signals, but not synaptic activity
Correlated network events in the developing neocortex were first reported by Rafael Yuste and Larry Katz in 1992 (Yuste et al., 1992). Using calcium imaging with the chemical indicator Fura2 in acute cortical slices from postnatal day (P) 0-7 rats, they described spontaneous events of correlated calcium fluctuations, which they referred to as neuronal domains. These domains engaged small cortical regions (< 100 μm in diameter) and were relatively infrequent (one event every 4 min; Fig. 1A). Those calcium transients were insensitive to the sodium channel blocker tetrodotoxin (TTX), and therefore not mediated by synaptic activity. Instead, they found that neuronal domains represented the propagation of calcium signals across gap junctions between neighboring neurons (see section 4.1). Thus, even though neuronal domains are often considered to be examples of synchronous developmental neuronal activity, they actually reflect correlated calcium events observed in acute cortical slices and independent of action potential firing. A related network phenomenon observed in vitro that is not driven by synaptic activity, synchronous plateau assemblies (SPAs), was first reported in the neonatal rat hippocampus after being triggered by the hormone oxytocin released by the mother during delivery (Crepel et al., 2007). SPAs were later described in neocortical neurons, as synchronous calcium waves observed in roughly half of acute cortical slices from P0-P3 mice (Allene et al., 2008). Like neuronal assemblies, SPAs are also gap-junction mediated, at least in the hippocampus (Crepel et al., 2007). To our knowledge, activity akin to neuronal domains or SPAs has only been observed in vitro, never in the intact brain. The significance of neuronal domains and their potential role in cortical development remain unclear to this day.
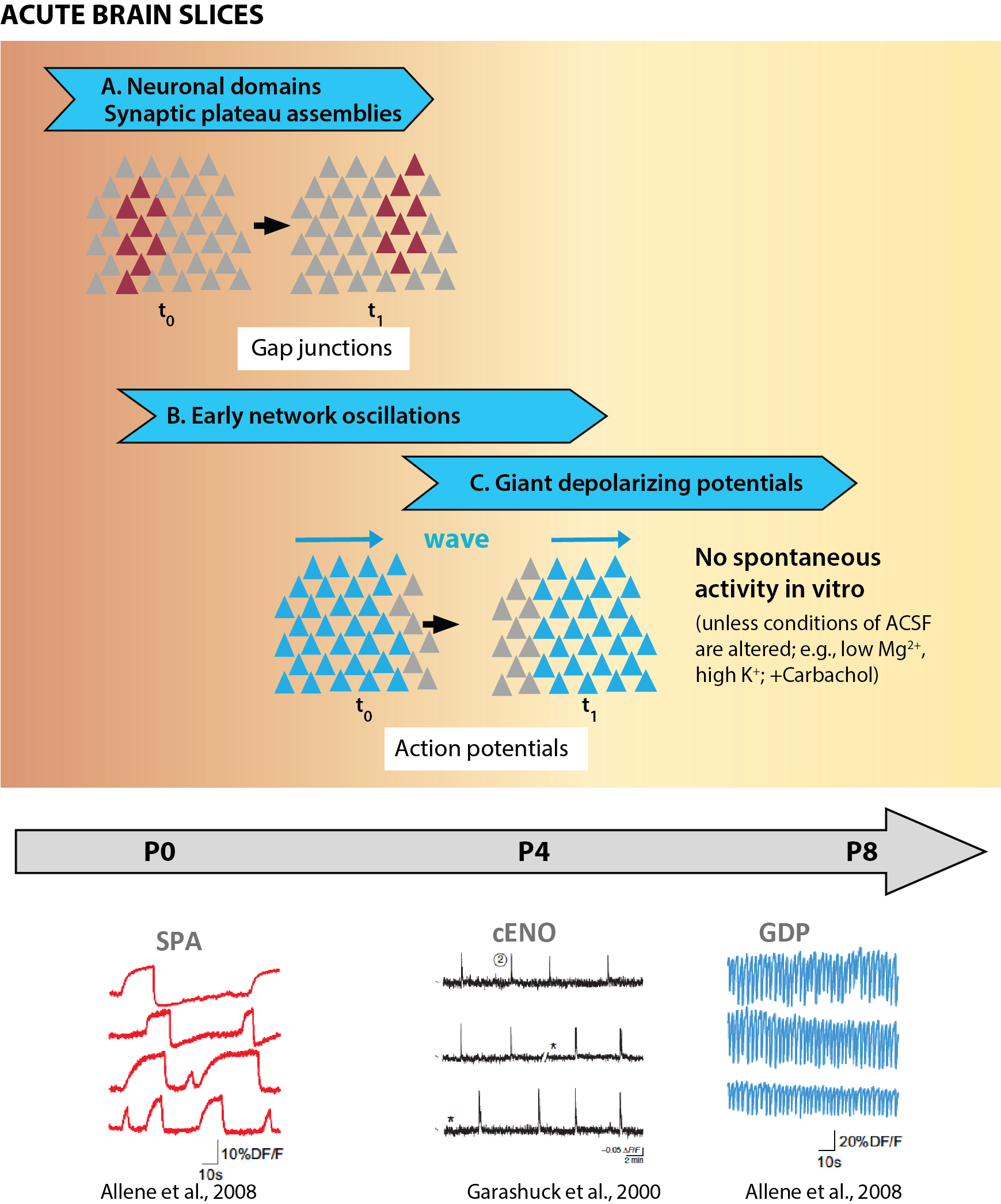
Fig 1 Spontaneous network activity in acute slices. (References: Allene et al. 2008, Garaschuk et al. 2000)
¶ 3. Early network oscillations revealed by calcium imaging in acute brain slices:
The first demonstration of activity-dependent correlated network events in cortical neurons at early postnatal ages came a quarter century ago (Corlew et al., 2004; Garaschuk et al., 1998; Garaschuk et al., 2000; Leinekugel et al., 1997). Using calcium imaging with chemical indicators (Fluo-3, Fura2) in acute brain slices, these pioneering studies revealed synchronous network events, which one group referred to as early network oscillations (ENOs) (Fig. 1B). ENOs are dependent on action potentials, as they are blocked by TTX (Corlew et al., 2004; Garaschuk et al., 2000), and rely on both excitatory and inhibitory synaptic transmission (see below). Subsequent in vitro calcium imaging studies of mouse neocortical slices with faster acquisition rates and improved calcium sensors have added little to our understanding of these early network events. Some studies have claimed to identify putative pacemaker regions from which ENOs arise and subsequently propagate as waves to the entire cortex, including the lateral entorhinal cortex (Lischalk et al., 2009; Namiki et al., 2013) or the septum (Conhaim et al., 2010). Surprisingly, the site of initiation was different depending on whether synchronous events were observed in horizontal vs. coronal brain slices (Namiki et al., 2013), though this could be an artifact of the slice preparation and may not translate in vivo.
One notable calcium imaging study of early postnatal rat cortical slices proposed a developmental sequence of two distinct network events, as follows: ENOs are generated first and then are followed by giant depolarizing potentials (GDPs) (Allene et al., 2008) (Fig. 1C). GDPs had been originally discovered with electrophysiology in hippocampal slices as synchronous events driven by depolarizing GABAergic transmission (Ben-Ari et al., 1989). The main differences between ENOs and GDPs are in their spatiotemporal dynamics, with ENOs being longer (>5 s for ENOs vs. <2 s for GDPs) and much less frequent (0.01 Hz for ENOs vs. 0.1 Hz for GDPs). Moreover, whereas GDPs are blocked by the GABAA-receptor antagonist bicuculline, ENOs are not (Allene et al., 2008).
Unfortunately, using brain slices to investigate the emergence of network activity during development has certain limitations owing to the severe disruption of circuits. In this review we will discuss how CNA is closely modulated by peripheral inputs and brainstem neuromodulation, both of which are disrupted in slices. The concern is that network activity patterns observed in acute slices are not necessarily good indicators of what happens in the intact brain of living animals. For example, ENOs cannot be detected in acute brain slices from mice/rats older than P7 (Corlew et al., 2004; Garaschuk et al., 1998; Garaschuk et al., 2000), presumably because cortical neurons are massively deafferented. In contrast, neuronal activity persists in vivo in the second postnatal week and remains highly correlated (as discussed below). Experimental conditions for in vitro recordings can be artificially manipulated to increase neuronal excitability and maintain some degree of spontaneous network activity in slices – for instance, ENOs re-emerge in older slices exposed to TTX (McCabe et al., 2006) or carbachol (Peinado et al., 1993). Moreover, some patterns of spontaneous activity in cortical slices, such as the high-frequency oscillation of GDPs as described by Allène and colleagues (Allene et al., 2008), have not been detected in vivo. In the end, attempts to classify different patterns of correlated activity in neocortical slices in chronological order (SPAs → ENOs → GDPs; Fig. 1) have not quite translated to the in vivo condition. Considering how large-scale circuit connectivity is disrupted in slices (e.g., the absence of bottom-up inputs and brainstem neuromodulation), conclusions from studies of correlated activity of cortical neurons in vitro should be interpreted with caution.
¶ 4. ENOs in vivo
The first in vivo recordings of ENOs were achieved in newborn mice using 2-photon calcium imaging (2PCI) with OGB1 at P5 (Adelsberger et al., 2005). The field of view was small because imaging was done through a small fiberoptic implant (200 μm tip diameter), but synchronous firing of large ensembles of neurons could clearly be resolved, and events were strikingly similar to cortical ENOs previously described in vitro in terms of their temporal dynamics (Garaschuk et al., 1998). The events were very sensitive to anesthesia, appearing only in unanesthetized mice, which suggests that a minimum network excitability is required to generate this correlated activity. Since then, many studies have reported synchronous network events in early postnatal mice using calcium imaging, across different cortical areas and different cortical layers, with a significant degree of similarity across studies (Che et al., 2018; Duan et al., 2020; Golshani et al., 2009; Kummer et al., 2016; Maldonado et al., 2021; Mizuno et al., 2018; Nakazawa et al., 2020; Rochefort et al., 2009; Siegel et al., 2012). Interestingly, some of the early in vivo 2PCI studies described two different stages of synchronous network activity in L2/3 of mouse somatosensory cortex at different stages of development (Golshani et al., 2009). First, at around P5-P8, highly synchronous activity was observed in spatially restricted modules, such that the highest correlation coefficients were observed in neuron pairs located within 100 μm. Next, around P8-P11, much larger events of synchronous neuronal activity spreading over >200 μm were observed. This progression of network events is discussed in more detail below (see also Fig. 2).
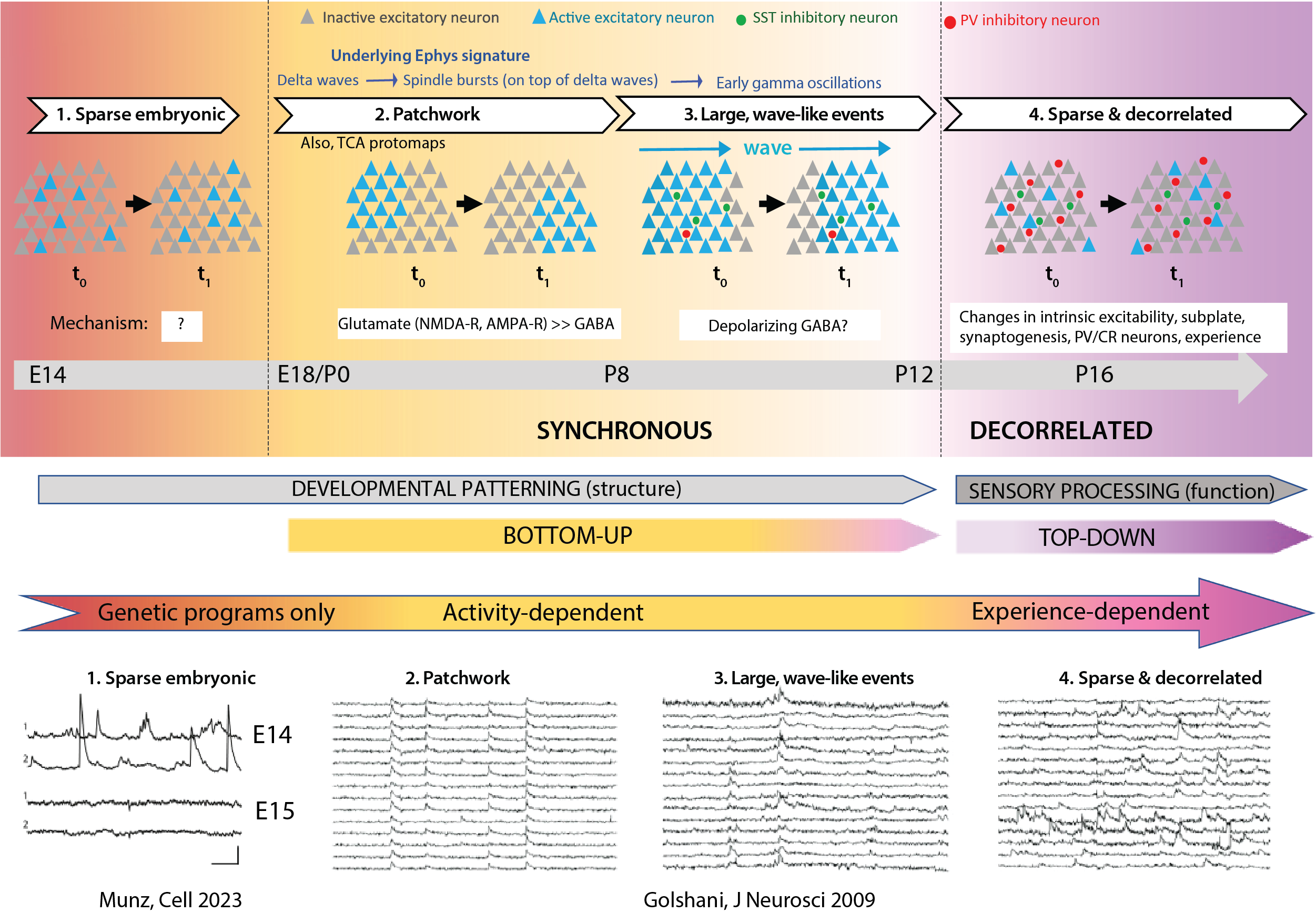
Fig 2 Spontaneous network activity in vivo. (References: Munz et al. 2023, Golshani et al. 2009)
¶ 5. Interneurons also participate in correlated network events:
The pioneering calcium imaging studies described above did not differentiate between excitatory and inhibitory neurons, as the chemical indicator dyes were taken up by both. Still, a few early studies suggested that interneurons also participate in CNA; for example, up to 70% of interneurons in acute hippocampal slices participate in ENOs (Garaschuk et al., 1998), and somatostatin (SST) interneurons help drive the synchronous activity (Flossmann et al., 2019; Picardo et al., 2011). More recent in vivo 2PCI experiments have revealed that both excitatory and inhibitory populations are active together in CNA (Duan et al., 2020; Kourdougli et al., 2023; Leighton et al., 2021). Although using Cre driver lines makes it possible to record from specific subclasses of interneurons, so far very few studies have recorded network dynamics of inhibitory neurons during early development. Therefore, the specific contributions of different subclasses of inhibitory neurons in CNA have only recently begun to be revealed.
After birth, GABAergic neurons derived from the medial and caudal ganglionic eminences (MGE, CGE, respectively) begin to populate the neocortex (BOX 2). Using in vivo calcium imaging in GAD67-Cre mice to record from all cortical interneurons, one study described their synchronous activity as early as P4-P6 (Modol et al., 2020). These GABAergic calcium events (GCEs) are rather complex phenomena, involving non-overlapping groups (or ‘assemblies’) of GAD67 neurons of various sizes. Interneurons of all major subclasses manifest similar GCEs (Modol et al., 2020), but with different characteristics. For example, future parvalbumin (PV) neurons within the MGE-derived population (in Lhx6-Cre mice) tend to segregate into functionally and spatially clustered assemblies to a greater extent than do CGE-derived interneurons (in 5HT3a-Cre mice) (Modol et al., 2020). In contrast, SST interneurons (recorded in SST-Cre mice) do not form assemblies and the frequency of their synchronous events is lower (Modol et al., 2020). Of note, at P4-P6, GCEs occur more frequently than global network events (i.e., events involving both glutamatergic and GABAergic neurons), whereas by P10-11, global network events take over, suggesting that interneurons may be the initial drivers of correlated activity. Future studies will be needed to confirm these results and further elucidate how interactions between GCEs of different classes of inhibitory neurons interact with CNA of excitatory neurons to shape the emergence of mature circuit dynamics.
¶ 6. Spindle bursts revealed by electrophysiology
Since CNA revealed by calcium imaging reflects neuronal action potential firing, what does it look like when recorded by electrophysiology? Khazipov, Buzsáki and colleagues were the first to use electrophysiology to record cortical network activity in vivo in the neonatal rodent brain, first in the hippocampus (Leinekugel et al., 2002), then in neocortex (Khazipov et al., 2004). In the primary somatosensory cortex (S1) of unanesthetized newborn rats, they observed neonatal network activity as spindle bursts, brief (lasting <1 s) spindle-shaped oscillations in the theta range (4-8 Hz) that are nestled within a delta wave and occur infrequently (separated by several seconds-long periods of silence) (Fig. 3A). Nearly identical spindle bursts were later observed in primary visual cortex (V1) (Colonnese et al., 2010; Hanganu et al., 2006; Hanganu et al., 2007). It is noteworthy that cortical neurons in culture show synchronous bursting when recorded by electrophysiology (Johnson and Buonomano, 2007; Maeda et al., 1995; Mohajerani and Cherubini, 2005). This suggests that synchronous bursting activity of cortical neurons reflects inherent network immaturity. The fact that spontaneous activity in the neonatal rodent cortex manifests as bursts of synchronous firing suggests that the cortex is intrinsically capable of generating CNA in the absence of external input (more on this below in section 3).
Based on results from electrophysiological recordings one can distinguish between three different stages of network activity in neonatal rodents stemming from chemical synapses (Colonnese et al., 2010; Khazipov and Luhmann, 2006; Khazipov and Milh, 2018) (Fig. 3B). First, at P0-P2, neocortical activity is dominated by slow delta waves. This is followed at P3-P8 by the appearance of spindle bursts on top of the delta waves (lasting 0.5-3 secs every 10 secs). Finally, a third stage (P8-P11), which in V1 coincides with the emergence of light responsivity, is still characterized by spindle bursts but also by short bouts of early gamma oscillations (EGOs), as well as by long oscillations that presumably synchronize activity over larger cortical regions (Colonnese et al., 2010). The long-range synchrony of spindle bursts across both hemispheres increases gradually throughout the first postnatal week (Yang et al., 2009).
The similarities between CNA recorded with calcium imaging (e.g., ENOs) and electrophysiology (e.g., delta waves with spindle bursts) in neonatal cortical neurons had always intimated that they represent the same phenomenon. Eventually, studies that combined in vivo extracellular recordings with 2P calcium imaging did confirm that spindle bursts represent the electrophysiological correlates of synchronous calcium events in the developing neocortex (Kirmse et al., 2015; Kummer et al., 2016) (Fig. 3C).
¶ 7. Activity patterns in the embryonic rodent neocortex
In recent years we have been learning more about activity patterns in the embryonic rodent brain in vivo. Documenting activity in vivo is now within our reach thanks to the latest methods being developed for imaging embryos in utero with cellular resolution (Huang et al., 2020; Munz et al., 2023; Yuryev et al., 2018). Using a remarkably ingenious approach for para-uterine calcium imaging of individual embryos in custom holders, it was possible to record from cortical neurons as early as embryonic day (E) 13.5 (Munz et al., 2023). This revealed that cortical activity of Rbp4+ neurons (future L5 excitatory neurons) in the embryonic brain emerges in two phases: a transient phase of sparse activity at E14.5 followed by two days of little or no spontaneous activity, and then a second more stable phase of activity after E17.5 (Munz et al., 2023). Interestingly, the pattern of activity at E18.5 was synchronous across different neuronal somata. This matches well the results of earlier studies that used transcranial calcium imaging in mouse embryo brains, which showed that spontaneous activity in S1 consists of distinct patches of correlated neuronal activity (Anton-Bolanos et al., 2019). Thus, CNA seems to be present during late embryonic development in mice, just before birth. Whether the earlier activity at E14.5 in vivo requires gap junctions (like neuronal domains in vitro), is not known.
¶ 8. Developmental sequence of events:
We have summarized above the timeline of discoveries concerning CNA in neocortex. Taking into consideration all the available data in this chronological account (which comes almost entirely from rodents and primarily from S1), we can now infer a developmental sequence of neuronal in the developing neocortex (Fig. 2). Compared to previous reviews on the topic, we have chosen not to include in this sequence activity patterns that have been reported in acute brain slices but not confirmed in vivo. Although we have focused on evidence from rodents to generate this sequence, it is important to note that CNA is highly conserved across species, and the development of cortical network activity probably follows similar stages in higher mammals.
¶ Stage 1
The earliest pattern of activity in the embryonic brain, seen at E14.5, is sparse and non-synchronous (Munz et al., 2023). Additional in vivo studies will be necessary to determine whether this scattered activity of individual neurons in the embryonic brain is dependent on gap junctions and therefore similar to neuronal domains (i.e., SPAs by electrophysiology). At present, the mechanisms and role of this activity remain a mystery.
¶ Stage 2
Beginning at E17.5-E18.5, synchronous network events become apparent and last until P6-P7. At this stage, and throughout the first postnatal days, this CNA is patchy, corresponding to individual cortical columns (or barrels in vS1), appearing and disappearing over time scales of tens of seconds. Stimulation of the whisker pad at E18.5 triggers local calcium events (Anton-Bolanos et al., 2019) that are very reminiscent of the patchwork activity seen in cortical L4 during the first postnatal days (Nakazawa et al., 2020), suggesting that these clusters represent functional protomaps that may correspond to synchronous firing of thalamocortical axons. In other words, it is conceivable that passive whisker movements in utero generate peripheral inputs that provide a template for shaping protomaps in L4. Around the end of the first postnatal week, patches of synchronous neurons about the size of an individual barrel become even more apparent in L2/3 (Golshani et al., 2009; Mizuno et al., 2018). At the electrophysiological level, this pattern of activity manifests as delta waves and spindle bursts (Khazipov et al., 2004; Minlebaev et al., 2011). These patches are mediated by glutamatergic synapses. Borrowing from Mizuno et al., we suggest that this stage be referred to as “patchwork activity”. We propose that the term ENO be reserved for describing similar events in vitro.
¶ Stage 3
In the second postnatal week, this patchwork pattern of CNA is replaced by larger events that propagate as waves and synchronize large swaths of neocortex (Golshani et al., 2009; Mizuno et al., 2018), marking the third stage of network activity. An appropriate term for this pattern of CNA activity would be “wave-like activity”. Individual neurons participating in these events still manifest spindle bursts (and early gamma oscillations), which reflects an increasing role of GABAergic synapses. Interestingly, the activity of inhibitory interneurons in S1 also appears to undergo a similar transition from small barrel-sized patches to larger wave-like events. Indeed, the size of GCEs more than doubles in size between P4-6 and P7-9, from ~150 μm in diameter (i.e., corresponding to the size of a single barrel) to 374 μm (Modol et al., 2020). The early interneuron assemblies at P4-6 are anatomically clustered and non-overlapping. After P10-11, separable assemblies are no longer observed. The increase in the number of GABAergic cell assemblies between P4-6 and P7-9, followed by the disappearance of distinct clusters, suggests that assembly size first matches that of single barrel, then expands across multiple barrels (Modol et al., 2020).
¶ Stage 4
By the end of the 2nd postnatal week, activity becomes decorrelated (Golshani et al., 2009; Rochefort et al., 2009). We propose the term “sparse network activity” (SNA) to describe this final mature stage of network activity (see below). With the maturation of both glutamatergic neurotransmission and the emergence of GABAergic inhibition within cortical circuits, the spontaneous activity of cortical neurons becomes decorrelated, whereby the proportion of cells participating in a given spontaneous network event is very low (Golshani et al., 2009; Rochefort et al., 2009). The exact mechanisms are not known (see below), but this developmental desynchronization might be responsible for the rapid refinement of sensory-evoked responses (Dorrn et al., 2010; Ikezoe et al., 2012; Stern et al., 2001). Furthermore, sparse activity of neurons is critical for coding information and is more efficient from an energy expenditure standpoint.